Pakar perubatan artikel itu

Penerbitan baru
Model eksperimen osteoarthritis
Ulasan terakhir: 07.07.2025

Semua kandungan iLive disemak secara perubatan atau fakta diperiksa untuk memastikan ketepatan faktual sebanyak mungkin.
Kami mempunyai garis panduan sumber yang ketat dan hanya memautkan ke tapak media yang bereputasi, institusi penyelidikan akademik dan, apabila mungkin, dikaji semula kajian secara medis. Perhatikan bahawa nombor dalam kurungan ([1], [2], dan lain-lain) boleh diklik pautan ke kajian ini.
Jika anda merasakan bahawa mana-mana kandungan kami tidak tepat, ketinggalan zaman, atau tidak dipersoalkan, sila pilih dan tekan Ctrl + Enter.
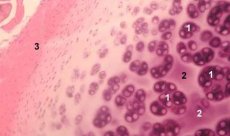
Rawan adalah tisu yang sangat khusus yang mengandungi hanya satu jenis sel (kondrosit) dan dicirikan oleh ketiadaan darah dan saluran limfa. Rawan terutamanya dipelihara oleh penyerapan daripada cecair sinovial. Metabolisme kondrosit dikawal oleh beberapa faktor larut yang dihasilkan secara tempatan oleh kondrosit dan tisu sekeliling. Fungsi Chondrocyte juga bergantung pada komposisi persekitaran ekstraselular (ketegangan oksigen, kepekatan ion, pH, dll.), komposisi ECM, interaksi sel dan matriks, dan isyarat fizikal. Objektif utama pemodelan eksperimen adalah untuk mencipta budaya dalam persekitaran ekstraselular tanpa mengubah fenotip sel matang. Objektif kedua adalah untuk mencipta budaya untuk mengkaji tindak balas pramatang, tertunda, jangka pendek, atau berpanjangan kondrosit kepada isyarat kimia dan/atau fizikal. Kajian in vitro juga memberi peluang untuk mengkaji tingkah laku kondrosit dalam osteoarthrosis. Objektif ketiga adalah untuk membangunkan sistem kultur yang membolehkan mengkaji interaksi pelbagai tisu dalam sendi. Tugas keempat ialah penyediaan implan tulang rawan untuk pemindahan seterusnya. Dan akhirnya, tugas kelima ialah kajian faktor pertumbuhan, sitokin atau agen terapeutik yang mampu merangsang pemulihan dan/atau menghalang penyerapan rawan.
Sepanjang dekad yang lalu, pelbagai model kultur sel rawan artikular telah dicipta, termasuk kultur monolayer, kultur terampai, kultur kondron, eksplan, kultur dan kultur sel abadi. Setiap budaya mempunyai kelebihan dan kekurangannya sendiri, dan masing-masing sesuai untuk mengkaji satu aspek khusus metabolisme kondrosit. Oleh itu, eksplan tulang rawan adalah model yang sangat baik untuk mengkaji perolehan unsur matriks, yang memerlukan reseptor permukaan sel tulen dan interaksi sel-matriks dan sel matriks yang normal. Pada masa yang sama, adalah disyorkan untuk mengkaji deposit matriks atau mekanisme yang mengawal metabolisme kondrosit pada budaya sel terpencil. Budaya monolayer berketumpatan rendah diperlukan untuk mengkaji proses pembezaan sel. Kultur terampai dalam matriks semula jadi atau sintetik adalah model untuk menganalisis tindak balas penyesuaian kondrosit kepada tekanan mekanikal.
Kultur kondrosit
Beberapa perkara penting harus diambil kira apabila memilih tisu rawan untuk kajian in vitro. Komposisi matriks dan aktiviti metabolik kondrosit berbeza-beza di antara sendi, dan yang terakhir juga bergantung pada kedalaman lokasi kondrosit dalam tisu. Data ini diperolehi dalam beberapa eksperimen di mana subpopulasi kondrosit terpencil dari zon rawan dengan kedalaman yang berbeza telah dikaji. Sejumlah perbezaan morfologi dan biokimia ditemui antara kondrosit berbudaya yang terletak di lapisan superfisial dan dalam rawan artikular. Sel dangkal mensintesis matriks fibrillar yang jarang, proteoglycan-miskin, manakala sel yang lebih dalam menghasilkan matriks yang kaya dengan fibril dan proteoglikan. Lebih-lebih lagi, sel-sel dangkal menghasilkan proteoglycan dan asid hyaluronik yang agak kecil dan agak kurang aggrecan dan keratan sulfat daripada kondrosit yang lebih dalam. Satu lagi ciri tersendiri penting bagi metabolisme kondrosit yang diasingkan daripada zon rawan dengan kedalaman yang berbeza ialah tindak balas kepada rangsangan eksogen. Menurut M. Aydelotte et al., kondrosit lembu dari zon rawan dangkal lebih sensitif terhadap IL-1 daripada sel dari zon dalam.
Tingkah laku sel juga bergantung pada lokasi tisu. Kondrosit daripada tulang rusuk dan tulang rawan telinga daripada haiwan yang sama bertindak balas secara berbeza kepada faktor pertumbuhan seperti faktor pertumbuhan fibroblast (FGF) dan TGF-beta. FGF meningkatkan penggabungan timidin, prolin dan leucine ke dalam tulang rusuk yang dikultur tetapi bukan kondrosit telinga. TGF-beta meningkatkan penggabungan timidin ke dalam kondrosit tulang rusuk dan rawan telinga tetapi tidak mempunyai kesan ke atas penggabungan timidin dan prolin ke dalam kondrosit telinga. Sel-sel rawan dari kawasan tekanan tinggi berbeza daripada sel-sel dari kawasan tekanan rendah pada rawan. Oleh itu, kondrosit tulang rawan sendi lutut biri-biri matang dari kawasan tengah permukaan artikular tibia yang tidak diliputi oleh meniskus, yang menanggung beban terbesar dalam vivo, mensintesis kurang aggrecan, tetapi lebih banyak hiasan daripada sel-sel dari zon yang diliputi oleh meniskus. Penulis juga menekankan kepentingan menggunakan rawan dari zon sendi yang sama apabila mengkaji fungsi sintetik sendi.
Metabolisme kondrosit dan tindak balas mereka terhadap faktor pengawalseliaan juga sangat bergantung pada usia penderma, perkembangan rangka mereka, dan keadaan sendi dari mana sel diambil. Dalam kondrosit manusia, penurunan ketara dalam tindak balas proliferatif dengan usia diperhatikan. Penurunan terbesar diperhatikan pada penderma berumur 40-50 tahun dan lebih 60 tahun. Selain itu, keterukan tindak balas proliferatif terhadap faktor pertumbuhan (cth, FGF dan TGF-beta) berkurangan dengan penuaan. Sebagai tambahan kepada perubahan kuantitatif dalam percambahan kondrosit, terdapat juga perubahan kualitatif. Sel-sel daripada penderma muda (10-20 tahun) bertindak balas lebih baik kepada faktor pertumbuhan terbitan platelet (PDGF) daripada TGF-beta, manakala sebaliknya diperhatikan dalam sel daripada penderma dewasa. Beberapa mekanisme digunakan untuk menerangkan perubahan yang bergantung kepada umur dalam fungsi sintetik kondrosit dan tindak balasnya terhadap faktor pertumbuhan. Ini termasuk penurunan bilangan dan pertalian reseptor sel permukaan, perubahan dalam sintesis dan bioaktiviti faktor pertumbuhan dan sitokin, dan pengubahsuaian isyarat pasca reseptor.
Keadaan patologi sendi juga mengubah morfologi dan aktiviti metabolik kondrosit. Oleh itu, J. Kouri et al. (1996) mengenal pasti tiga subpopulasi kondrosit dalam rawan dalam osteoarthrosis. Kondrosit dari bahagian cetek dan atas bahagian tengah rawan membentuk kelompok dan mensintesis lebih banyak proteoglikan dan kolagen. TGF-beta dan faktor pertumbuhan seperti insulin (IGF) mampu merangsang sintesis proteoglikan oleh kondrosit dan meneutralkan sebahagian kesan IL-1 dan TNF-a. Eksplan tulang rawan yang terjejas oleh osteoarthrosis dan kondrosit yang diasingkan daripada rawan pesakit dengan osteoarthrosis adalah lebih sensitif terhadap rangsangan TGF-beta daripada kondrosit rawan yang sihat. Perbezaan ini berkemungkinan besar dikaitkan dengan perubahan fenotip dalam kondrosit pada lapisan atas rawan artikular.
Pengasingan kondrosit individu dicapai dengan rawatan berurutan ECM dengan enzim proteolitik. Selepas pembebasan mereka daripada ECM, sel terpencil adalah sesuai untuk mengkaji sintesis de novo komponen matriks. Sesetengah pengarang hanya menggunakan kolagenase clostridial, manakala yang lain pra-inkubasi rawan dengan trypsin, pronase, DNase dan/atau hyaluronidase. Bilangan sel terpencil bergantung kepada enzim yang digunakan. Oleh itu, apabila dirawat dengan kolagenase sahaja, 1.4-10 6 kondrosit boleh diperoleh daripada 1 g tisu, sambil menggunakan pronase, hyaluronidase, dan kolagenase - 4.3-10 6. Apabila dirawat dengan kolagenase, aggrecan, protein, IL-6, dan IL-8 kekal dalam kultur sel dalam kuantiti yang jauh lebih besar daripada rawatan berurutan dengan enzim yang berbeza. Terdapat beberapa penjelasan untuk perbezaan ini antara kedua-dua kultur sel:
- Reseptor sel rosak atau dihalang oleh enzim, TGF-beta menghalang sintesis DNA dan proteoglycan dalam kondrosit yang baru diasingkan (hari 1), manakala sintesis DNA dan proteoglycan dalam kondrosit yang dibiakkan dalam monolayer (7 hari) dirangsang oleh TGF-beta. Walau bagaimanapun, tempoh yang mencukupi diperlukan untuk ekspresi semula komponen membran ini sebelum permulaan eksperimen.
- Protease eksogen boleh mengganggu interaksi sel-matriks pengantara integrin. Keluarga integrin menggalakkan perlekatan kondrosit pada molekul ECM (Shakibaei M. et al., 1997). Gangguan ini boleh menjejaskan ekspresi gen matriks.
- Sisa komponen matriks boleh mengawal fungsi sintetik kondrosit. Integrins dapat mengenali produk degradasi ECM, dengan itu memainkan peranan penting dalam pembaikan tisu selepas tindakan enzim proteolitik. T. Larsson et al. (1989) melaporkan bahawa penambahan proteoglikan utuh atau berpecah kepada kultur sel merangsang sintesis protein dan proteoglikan. Walau bagaimanapun, tahap asid hyaluronik yang tinggi menyebabkan penurunan ketara dalam kemasukan sulfat dalam sintesis proteoglikan oleh kondrosit embrio ayam, kondrosit matang sel chondrosarcoma babi dan tikus. Selain itu, asid hyaluronik adalah perencat pembebasan proteoglikan daripada sel walaupun dengan kehadiran IL-1b, TNF-a, FGF, yang menunjukkan tindak balas aktiviti biologi pertama faktor pertumbuhan dan sitokin. Mekanisme tepat yang mendasari tindakan asid hyaluronik masih tidak jelas; Chondrocytes diketahui mengandungi reseptor untuk asid hyaluronik yang dikaitkan dengan filamen aktin sitosol. Pengikatan asid hyaluronik pada reseptornya merangsang fosforilasi protein. Oleh itu, data ini menunjukkan modulasi fungsi metabolik kondrosit oleh molekul protein matriks yang berpecah atau asli melalui pengaktifan reseptor membran sel.
- Rangsangan pantas sintesis protein matriks oleh kondrosit oleh enzim mungkin disebabkan oleh perubahan dalam bentuk kondrosit dan/atau penyusunan semula sitoskeletal.
- Sesetengah sitokin (cth, IL-8) dan faktor pertumbuhan (cth, IGF-1, TGF-β) diasingkan dalam ECM. Contoh yang paling terkenal ialah pengikatan TGF-β oleh decorin, yang mengakibatkan penurunan keupayaan bekas untuk mendorong pertumbuhan sel dalam sel ovari hamster Cina. Penemuan bahawa kandungan decorin dalam rawan meningkat dengan usia menunjukkan penurunan dalam bioavailabiliti TGF-β dengan penuaan. Faktor pertumbuhan dan sitokin boleh dibebaskan daripada serpihan matriks semasa kultur dan seterusnya memodulasi fungsi kondrosit.
Budaya monolayer kondrosit
Fenotip kondrosit yang dibezakan terutamanya dicirikan oleh sintesis kolagen jenis II dan proteoglycans khusus tisu, serta tahap aktiviti mitosis yang rendah. Terdapat bukti bahawa dengan penanaman sel yang berpanjangan dalam lapisan tunggal, serta selepas beberapa laluan sel berulang, kondrosit kehilangan garis sferanya dan memperoleh bentuk seperti fibroblas yang memanjang. Dengan metaplasia fibroblastik sedemikian, fungsi sintetik sel juga diubah suai, dicirikan oleh penurunan progresif dalam sintesis kolagen jenis II, IX dan XI dan peningkatan dalam sintesis kolagen jenis I, III dan Y. Proteoglikan kecil yang tidak terkumpul disintesis disebabkan oleh aggrecan berfungsi. Sintesis cathepsin B dan L sangat rendah dalam sel yang dibezakan, tetapi meningkat dalam proses kehilangan pembezaan. Kolagenase-1 dinyatakan dalam kondrosit yang dibezakan; dengan penanaman yang berpanjangan, ekspresinya berkurangan, manakala pengeluaran perencat tisu metalloproteases (TIMPs) meningkat.
Kondrosit yang dibezakan mengekspresikan semula kolagen bagi fenotip yang dibezakan apabila dipindahkan daripada lapisan tunggal kepada budaya terampai. Proses pembezaan mungkin berkaitan dengan bentuk sel. Harta ini kerap digunakan oleh penyelidik yang mengkaji cantuman yang rosak dengan kondrosit autologous. Sebilangan kecil sel yang diperoleh daripada bahan biopsi boleh dikembangkan dalam budaya monolayer dan kemudian dimasukkan semula ke dalam matriks tiga dimensi sebelum pemindahan. Ekspresi semula fenotip tertentu oleh kondrosit terpebeza yang dipindahkan ke kultur agarose boleh dirangsang oleh TGF-β, kompleks ossein-hydroxyapatite, dan asid askorbik.
Sebagai tindak balas kepada faktor pertumbuhan dan sitokin, kondrosit diubah suai semasa proses pembezaan. Tindak balas selular terhadap sitokin dan faktor pertumbuhan berbeza antara kondrosit yang tidak dibezakan dan dibezakan. IL-1 merangsang percambahan fibroblast, manakala pertumbuhan kondrosit yang tidak dibezakan dihalang oleh IL-1. Sintesis DNA dirangsang oleh IGF-1 dalam kondrosit yang memanjang tetapi tidak diratakan. Dalam kondrosit yang dibezakan, kesan rangsangan IL-1beta dan TNF-a pada pengeluaran procollagenase adalah lebih ketara berbanding dengan kondrosit yang tidak dibezakan.
Penanaman Kondrosit
Penanaman kondrosit dalam ampaian dalam medium cecair atau dalam matriks tiga dimensi semulajadi atau sintetik menstabilkan fenotip kondrosit. Sel-sel mengekalkan bentuk sferanya dan mensintesis protein khusus tisu. Kultur kondrosit yang digantung biasanya disyorkan untuk mengkaji pembentukan matriks perisel baru. Kultur kondrosit dalam polimer penyerap sintetik atau semulajadi digunakan untuk implantasi sel ke dalam kecacatan rawan untuk merangsang pertumbuhan semula tisu rawan sendi. Medium sintetik atau semulajadi untuk sel implan mesti memenuhi beberapa keperluan:
- implan mesti mempunyai struktur berliang untuk melekat dan pertumbuhan sel,
- polimer itu sendiri atau produk degradasinya tidak boleh menyebabkan keradangan atau tindak balas toksik apabila ditanam dalam vivo,
- pembawa cantuman mesti mempunyai keupayaan untuk mengikat rawan bersebelahan atau tulang subkondral,
- matriks semula jadi atau sintetik mesti mempunyai keupayaan untuk menyerap, degradasinya mesti diseimbangkan oleh penjanaan semula tisu,
- untuk memudahkan pembaikan rawan, struktur kimia dan seni bina liang matriks mesti memudahkan penyelenggaraan fenotip selular dan sintesis protein khusus tisu oleh kondrosit yang diletakkan di dalamnya,
- Semasa implantasi in vivo, adalah perlu untuk mengkaji sifat mekanikal matriks sintetik atau semula jadi.
 [ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
[ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
Penggantungan kondrosit dalam fasa cecair
Lekatan sel pada salur plastik di mana kondrosit dikultur boleh dihalang dengan menyalut dindingnya dengan larutan metilselulosa, agarose, hidrogel (poli-2-hidroksietil metakrilat), atau campuran kolagen-agarose. Di bawah keadaan ini, kondrosit membentuk kelompok dan mensintesis terutamanya aggrecan dan kolagen khusus tisu (jenis II, IX, XI). Dua jenis sel biasanya dijumpai. Sel-sel yang terletak di tengah mengekalkan bentuk sfera dan dikelilingi oleh ECM yang dibangunkan dengan baik, yang disahkan oleh kajian histokimia dan ultrastruktur. Di pinggir, kondrosit mempunyai garis besar diskoid dan dikelilingi oleh ECM yang jarang; sedikit yang diketahui tentang ciri fungsi sel tersebut.
Adalah mungkin untuk memupuk kondrosit pada pembawa mikro yang dikekalkan dalam penggantungan; manik dextran (cytodex), manik dextran bersalut kolagen (cytodex III), dan mikrosfera tidak berliang jenis I kolagen (cellagen) digunakan sebagai pembawa mikro. Di bawah keadaan kultur ini, kondrosit melekat pada permukaan pembawa mikro, mengekalkan bentuk sferanya, dan menghasilkan bahan seperti matriks. Selain itu, penggunaan cellagen menggalakkan percambahan kondrosit dan ekspresi semula fenotip biasa. Oleh itu, pengkulturan kondrosit pada mikrosfera cellagen boleh digunakan untuk memulihkan fenotip sel sebelum pemindahan.
Kaedah lain untuk mengkultur suspensi kondrosit dalam medium cecair ialah penanaman mereka dalam bentuk bola padat yang terdiri daripada sel (0.5-1 * 10 b ), diperoleh dengan sentrifugasi. Kondrosit sedemikian mampu menghasilkan matriks yang mengandungi sejumlah besar proteoglycans, kolagen jenis II, tetapi bukan kolagen jenis I, yang disahkan oleh kaedah histologi, imunohistokimia dan kuantitatif.
Penggantungan kondrosit dalam ECM semulajadi
Kondrosit boleh dibiakkan dalam ampaian dalam matriks tiga dimensi (agar lembut, agarose, gel atau span kolagen, asid hyaluronik, gam fibrin, manik alginat).
Kondrosit yang dikultur dalam agarose mengekalkan fenotip normalnya dan mensintesis kolagen jenis II dan agregat agregat khusus tisu. Apabila dikultur dalam agarose, proteoglikan yang disintesis oleh sel dilepaskan ke dalam medium selama 50 hari. Sebagai perbandingan, dalam budaya monolayer, fasa selular dipenuhi dengan glikosaminoglikan dalam 5-6 hari pertama penanaman; apabila dibiakkan dalam medium, selepas peningkatan sintesis dan pembebasan glikosaminoglikan dalam 8-10 hari pertama, penurunan bergantung pada masa mereka berlaku. Walau bagaimanapun, tingkah laku kondrosit apabila dibiakkan dalam agarose berbeza daripada dalam vivo. Dalam agarose, sebilangan besar agregat agregat yang disintesis mengandungi molekul yang lebih kecil dan lebih sedikit molekul daripada dalam vivo. TGF-β merangsang sintesis proteoglycan dalam eksplan, tetapi mengurangkan sintesis aggrecan dalam agarose.
Alginat ialah polisakarida linear yang diperoleh daripada rumpai laut coklat. Dengan adanya kation divalen, seperti ion Ca 2+, polimer ini menjadi gel. Setiap kondrosit yang terperangkap dalam alginat dikelilingi oleh matriks polisakarida bercas negatif, yang liangnya setanding dengan yang terdapat dalam rawan hialin. Matriks yang dibentuk oleh kondrosit dalam manik alginat terdiri daripada dua petak - lapisan nipis matriks berkaitan sel yang sepadan dengan matriks perisel dan wilayah rawan artikular dan matriks yang lebih jauh bersamaan dengan antara wilayah dalam tisu asli. Pada hari ke-30 kultur, isipadu relatif dan mutlak yang diduduki oleh sel dan setiap dua petak dalam manik alginat adalah hampir sama sepenuhnya dengan yang terdapat dalam rawan asli. Selama hampir 30 hari, kondrosit mengekalkan bentuk sferanya dan menghasilkan aggrecan, sifat hidrodinamik yang serupa dengan molekul aggrecan dalam matriks rawan artikular, serta molekul kolagen jenis II, IX, dan XI. Pada masa yang sama, seperti kultur ampaian yang lain, sel yang diratakan terdapat pada permukaan manik alginat, yang menghasilkan sejumlah kecil molekul kolagen jenis I, yang dilepaskan terus ke dalam medium dan tidak dimasukkan ke dalam ECM. Pembiakan sederhana kondrosit diperhatikan dalam manik alginat. Selepas 8 bulan penanaman dalam gel alginat, kondrosit matang tidak kehilangan aktiviti metabolik dan terus mensintesis kolagen jenis II dan aggrecan khusus tisu.
H. Tanaka et al. (1984) menyiasat sifat resapan pelbagai molekul semula jadi dalam alginat dan mendapati bahawa molekul yang lebih besar daripada 70 kDa tidak meresap melalui alginat. Oleh itu, kultur sel dalam alginat sesuai untuk mengkaji peraturan biosintesis matriks dan organisasi ECM. Ketersediaan sel yang dikultur dalam alginat membolehkan seseorang mengkaji tindakan faktor pengawalseliaan peptida dan agen farmakologi pada tahap transkripsi, pasca-transkripsi, dan translasi.
Kondrosit juga dibiakkan dalam matriks gentian kolagen jenis I dan II. S. Nehrer et al. (1997) membandingkan fungsi kondrosit anjing dalam matriks polimer kolagen-proteoglikan berliang yang mengandungi kolagen daripada jenis yang berbeza. Mereka mendapati perbezaan penting dalam morfologi fungsi biosintetik kondrosit yang dikultur dalam matriks kolagen yang mengandungi jenis kolagen I dan II. Sel dalam matriks jenis kolagen II mengekalkan bentuk sferanya, manakala dalam jenis kolagen I ia mempunyai morfologi seperti fibroblas. Selain itu, dalam matriks jenis kolagen II, kondrosit menghasilkan jumlah glikosaminoglikan yang lebih besar. J. van Susante et al. (1995) membandingkan sifat-sifat kondrosit yang dikultur dalam gel alginat dan kolagen (jenis I). Penulis mendapati peningkatan ketara dalam bilangan sel dalam gel kolagen, tetapi dari hari ke-6 penanaman sel-sel kehilangan fenotip ciri mereka, bertukar menjadi sel seperti fibroblast. Dalam gel alginat, penurunan bilangan sel diperhatikan, tetapi kondrosit mengekalkan fenotip normalnya. Dalam gel kolagen, bilangan proteoglikan setiap sel adalah jauh lebih tinggi daripada alginat, tetapi dalam gel penurunan dalam sintesis unsur matriks diperhatikan, bermula dari hari ke-6 penanaman, manakala dalam alginat sintesis terus meningkat.
Matriks fibrin tiga dimensi pepejal ialah bahan semula jadi yang menyokong kondrosit yang terampai di dalamnya dalam fenotip terbeza. Matriks fibrin tiga dimensi juga boleh digunakan sebagai pembawa dalam pemindahan kondrosit. Kelebihan fibrin adalah ketiadaan sitotoksisiti, keupayaan untuk mengisi ruang, dan kapasiti pelekat. Kajian histologi dan biokimia, autoradiografi, dan mikroskop elektron telah menunjukkan bahawa kondrosit dalam gel fibrin mengekalkan morfologinya, membiak, dan menghasilkan matriks walaupun selepas 2 minggu penanaman. Walau bagaimanapun, G. Homminga et al. (1993) melaporkan bahawa pemecahan fibrin bermula selepas 3 hari penanaman, dan dediferensiasi kondrosit berkembang.
Penggantungan kondrosit dalam ECM buatan (sintetik).
Implan rawan untuk pembedahan rekonstruktif atau ortopedik boleh diperolehi dengan menumbuhkan kondrosit terpencil secara in vitro dalam matriks biokompatibel sintetik.
Kondrosit yang dikultur dalam asid polyglycolic membiak dan mengekalkan morfologi dan fenotip normal selama 8 minggu. Kompleks asid chondrocyte-polyglycolic terdiri daripada sel, glycosaminoglycans, kolagen, dan mempunyai kapsul kolagen luaran. Walau bagaimanapun, implan tersebut mengandungi dua jenis molekul kolagen - I dan II. Implan daripada kondrosit yang dibezakan oleh satu siri laluan mempunyai jumlah glikosaminoglikan dan kolagen yang lebih banyak daripada implan daripada kondrosit yang tidak dibezakan terutamanya.
L. Freed et al. (1993b) membandingkan tingkah laku kultur kondrosit manusia dan lembu dalam asid polyglycolic gentian (FPGA) dan asid polilaktik berliang (PPLA). Selepas 6-8 minggu mengkultur kondrosit lembu dalam FPGA atau PPLA, penulis memerhatikan percambahan sel dan penjanaan semula matriks rawan. Dalam FPGA, kondrosit mempunyai bentuk sfera dan terletak di lacunae yang dikelilingi oleh matriks rawan. Selepas 8 minggu kultur in vitro, tisu yang dijana semula mengandungi sehingga 50% bahan kering (4% jisim selular, 15% glikosaminoglikan, dan 31% kolagen). Dalam PPLA, sel mempunyai bentuk berbentuk gelendong dan sejumlah kecil glikosaminoglikan dan kolagen. Dalam FPGA, pertumbuhan sel adalah 2 kali lebih sengit daripada PPLA. Dalam vivo, kondrosit yang tumbuh dalam VPGK dan PPLC menghasilkan tisu secara histologi yang serupa dengan rawan dalam tempoh 1-6 bulan. Implan mengandungi glycosaminoglycans, kolagen jenis I dan II.
Kondrosit janin lembu telah dibiakkan dalam polietilena hidrofobik dan hidrofilik berketumpatan tinggi berliang. Selepas 7 hari pengeraman dalam kedua-dua substrat, sel-sel mengekalkan bentuk sfera dan mengandungi terutamanya kolagen jenis II. Selepas 21 hari penanaman, matriks hidrofilik didapati mengandungi lebih banyak kolagen jenis II daripada matriks hidrofobik.
Tisu rawan juga boleh diperolehi dengan mengkultur dalam satu lapisan pada penapis Millicell-CM. Pra-salutan penapis dengan kolagen adalah perlu untuk melekatkan kondroitin. Pemeriksaan histologi kultur menunjukkan pengumpulan kondrosit dalam ECM yang mengandungi proteoglikan dan kolagen jenis II. Kolagen jenis I tidak dikesan dalam budaya sedemikian. Kondrosit dalam tisu tulang rawan yang diperolehi adalah berbentuk sfera, tetapi pada permukaan tisu mereka agak rata. Ketebalan tisu yang baru terbentuk meningkat dengan masa dan bergantung kepada ketumpatan awal monolayer sel. Di bawah keadaan kultur optimum, ketebalan tisu rawan mencapai 110 μm, organisasi sel dan kolagennya ke dalam lapisan cetek dan dalam adalah serupa dengan rawan artikular. ECM mengandungi kira-kira 3 kali lebih banyak kolagen dan proteoglikan. Selepas 2 minggu penanaman, pengumpulan matriks diperhatikan, membolehkan tisu diekstrak daripada penapis dan digunakan untuk pemindahan.
Sims et al. (1996) mengkaji penanaman kondrosit dalam gel polietilena oksida, matriks polimer terkapsul yang membolehkan pemindahan sejumlah besar sel melalui suntikan. Enam minggu selepas suntikan ke dalam tisu subkutaneus tikus athymic, rawan baru terbentuk, yang secara morfologi dicirikan oleh opalescence putih yang serupa dengan rawan hialin. Data histologi dan biokimia menunjukkan kehadiran kondrosit yang membiak secara aktif menghasilkan ECM.
Penjelasan
Penjelasan tisu tulang rawan digunakan untuk mengkaji proses ana- dan katabolisme di dalamnya, penyelenggaraan homeostasis, penyerapan dan pembaikan. Kondrosit dalam eksplan tulang rawan mengekalkan fenotip normal dan komposisi ECM sama seperti dalam rawan artikular dalam vivo. Selepas 5 hari pengkulturan dengan kehadiran serum, tahap sintesis yang berterusan dan proses degradasi semula jadi dicapai. Penyerapan tisu boleh dipercepatkan dalam kultur utama dan dalam kultur dengan penambahan serum menggunakan beberapa agen, seperti IL-IB, TNF-a, lipopolisakarida bakteria, derivatif asid retinoik atau radikal oksigen aktif. Untuk mengkaji pembaikan rawan, kerosakannya disebabkan oleh mediator keradangan larut (H 2 O 2, IL-1, TNF-a) atau pecah fizikal matriks.
Kaedah kultur organotip adalah model untuk kajian in vitro tentang kesan faktor luaran terpencil pada kondrosit dan matriks sekeliling. Dalam vivo, kondrosit terletak jarang dalam ECM dan tidak bersentuhan antara satu sama lain. Budaya eksplan tulang rawan artikular mengekalkan organisasi struktur ini, serta interaksi khusus antara kondrosit dan persekitaran ekstraselular di sekelilingnya. Model ini juga digunakan untuk mengkaji kesan tekanan mekanikal, agen farmakologi, faktor pertumbuhan, sitokin, dan hormon pada metabolisme rawan.
Satu lagi kelebihan eksplanasi tisu tulang rawan ialah ketiadaan kerosakan pada kondrosit di bawah tindakan enzim proteolitik atau faktor mekanikal, yang tidak dapat dielakkan apabila mengasingkan sel. Reseptor dan protein membran dan glikoprotein lain dilindungi daripada faktor yang merosakkan.
[ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ]
Budaya Chondron
Chondron ialah unit struktur, berfungsi dan metabolik rawan artikular yang terdiri daripada kondrosit, matriks perisel dan kapsul filamen padat dan bertanggungjawab untuk homeostasis matriks. Kondron diekstrak secara mekanikal daripada rawan dan dikumpulkan menggunakan beberapa homogenisasi berkelajuan rendah berturut-turut. Kondron yang diasingkan daripada zon kedalaman rawan yang berbeza boleh dibahagikan kepada empat kategori: kondron tunggal, kondron berpasangan, kondron berbilang (tiga atau lebih) tersusun secara linear (lajur kondron), dan kelompok kondron.
Kondron tunggal biasanya ditemui di lapisan tengah rawan utuh, kondron berpasangan ditemui di sempadan lapisan tengah dan dalam, kondron berbilang yang tersusun secara linear adalah tipikal lapisan dalam rawan utuh. Akhir sekali, kelompok kondron terdiri daripada kumpulan kondron tunggal dan berpasangan yang teratur secara rawak, yang mengekalkan keadaan terkumpul selepas penghomogenan. Kelompok kondron ialah serpihan besar rawan, biasanya mengandungi beberapa kondron dan fibril kolagen tersusun jejari, iaitu, ciri organisasi tipikal lapisan dalam matriks. Kondron tidak bergerak dalam agarose telus, yang membolehkan kajian struktur, komposisi molekul, dan aktiviti metaboliknya. Sistem chondron-agarose dianggap sebagai mikromodel rawan, yang berbeza daripada sistem chondrocyte-agarose tradisional kerana persekitaran mikro semulajadi dipelihara, dan tidak perlu mensintesis dan memasangnya. Kultur Chondron ialah model untuk mengkaji interaksi sel dan matriks dalam rawan artikular di bawah keadaan normal dan patologi.
[ 22 ], [ 23 ], [ 24 ], [ 25 ], [ 26 ], [ 27 ]
Budaya kondrosit abadi
DNA rekombinan atau virus yang mengandungi onkogen yang mampu membuat sel "abadi" digunakan untuk mencipta garisan sel kekal. Kondrosit abadi mempunyai keupayaan untuk membiak tanpa henti sambil mengekalkan fenotip yang stabil. F. Mallein-Gerin et al. (1995) menunjukkan bahawa onkogen SV40T mendorong percambahan kondrosit tetikus, yang terus mengekspresikan jenis kolagen II, IX, dan XI secara stabil, serta protein aggrecan artikular dan mengikat. Walau bagaimanapun, garisan sel sedemikian memperoleh keupayaan untuk mensintesis kolagen jenis I apabila mengkulturnya dalam budaya monolayer atau dalam gel agarose.
W. Horton et al. (1988) menggambarkan barisan sel abadi dengan tahap ekspresi rendah mRNA kolagen jenis II. Sel-sel ini diperoleh dengan transformasi mereka dengan retrovirus tetikus yang mengandungi I-myc- dan y-ra-oncogenes. Sel jenis ini mewakili model unik untuk mengkaji interaksi matriks artikular tanpa adanya kolagen jenis II, serta peraturan sintesis kolagen jenis II.
Budaya kondroprit dengan gen bermutasi atau dipadam adalah model yang mudah untuk mengkaji fungsi fisiologi mereka. Model ini amat sesuai untuk mengkaji peranan molekul tertentu dalam organisasi matriks rawan atau untuk menyiasat kesan pelbagai faktor pengawalseliaan ke atas metabolisme rawan. Kondrosit dengan gen yang dipadamkan untuk kolagen jenis IX mensintesis fibril kolagen yang lebih lebar daripada biasa, menunjukkan bahawa kolagen jenis IX mengawal diameter fibril. Seperti yang dinyatakan dalam Bab 1, mutasi dalam pengekodan gen COLAI kolagen jenis II baru-baru ini telah ditemui dalam keluarga dengan osteoarthritis umum primer. Untuk mengkaji kesan kolagen jenis II mutan pada matriks artikular, R. Dharmrvaram et al. (1997) ditransfektifkan (dijangkiti asid nukleik asing) COL 2 AI yang rosak (arginine pada kedudukan 519 digantikan oleh sistein) ke dalam kondrosit janin manusia secara in vitro.
Sistem kebudayaan. Dalam sendi, rawan berinteraksi dengan jenis sel lain yang terkandung dalam membran sinovial, cecair sinovial, ligamen, dan tulang subkondral. Metabolisme kondrosit boleh dipengaruhi oleh pelbagai faktor larut yang disintesis oleh sel yang disenaraikan. Oleh itu, dalam arthritis, rawan artikular dimusnahkan oleh enzim proteolitik dan radikal bebas yang dihasilkan oleh sel sinovial. Oleh itu, model telah dibangunkan untuk mengkaji interaksi kompleks antara rawan dan tisu sekeliling, yang dipanggil cocultures.
S. Lacombe-Gleise et al. (1995) kultur kondrosit arnab dan osteoblas dalam sistem kokultur (COSTAR) di mana sel-sel dipisahkan oleh membran mikroporous (0.4 μm) yang membenarkan pertukaran antara kedua-dua jenis sel tanpa sebarang sentuhan langsung. Kajian ini menunjukkan keupayaan osteoblas untuk merangsang pertumbuhan kondrosit melalui mediator larut.
AM Malfait dan pengarang bersama (1994) mengkaji hubungan antara monosit darah periferi dan kondrosit. Model ini sesuai untuk mengkaji proses pengantara sitokin dalam arthropathies radang (artritis reumatoid, spondyloarthritides seronegatif, dll.). Pengarang model memisahkan sel dengan membran pengikat protein dengan liang diameter 0.4 μm. Kajian menunjukkan bahawa monosit yang dirangsang dengan lipopolisakarida menghasilkan IL-1 dan TNF-a, yang menghalang sintesis aggrecan oleh kondrosit dan menyumbang kepada degradasi agregat agregat yang telah disintesis.
K. Tada et al. (1994) mencipta model kokultur di mana sel endothelial dalam gel kolagen (jenis I) diletakkan di dalam ruang dalam yang dipisahkan dari ruang luar dengan kondrosit diletakkan di dalamnya oleh penapis dengan saiz liang 0.4 μm. Dalam keadaan pengasingan lengkap dari ruang luar, sel endothelial manusia membentuk tiub dalam gel kolagen dengan kehadiran EGF atau TGF-a. Apabila kedua-dua jenis sel dibiakkan secara serentak, pembentukan tiub yang bergantung kepada TGF oleh sel endothelial telah dihalang. Perencatan proses ini oleh kondrosit telah dihapuskan sebahagiannya oleh antibodi anti-TGF-beta. Ia boleh diandaikan bahawa TGF-beta yang dihasilkan oleh kondrosit menghalang vaskularisasi rawan itu sendiri.
S. Groot et al. (1994) secara serentak mengkultur kondrosit daripada zon hipertrofik dan proliferatif tulang janin tikus berusia 16 hari dengan kepingan tisu otak. Selepas 4 hari penanaman, transdifferentiation kondrosit kepada osteoblas dan permulaan pembentukan osteoid diperhatikan. Selepas 11 hari penanaman, sebahagian daripada rawan digantikan oleh tisu tulang dan matriks tulang telah dikalsifikasikan sebahagiannya. Sesetengah neuropeptida dan neurotransmitter yang dihasilkan oleh tisu otak menjejaskan metabolisme osteoblas atau mempunyai reseptor untuknya. Antaranya ialah norepinephrine, peptida usus vasoaktif, peptida berkaitan gen kalsitonin, bahan P dan somatostatin. Kepingan tisu otak yang dikultur bersama-sama dengan kondrosit boleh menghasilkan beberapa faktor tersenarai yang mampu mendorong proses transdifferentiasi kondrosit kepada osteoblas.
[ 28 ], [ 29 ], [ 30 ], [ 31 ], [ 32 ], [ 33 ]
Pengaruh faktor luaran pada budaya kondrosit
Pengaruh ketegangan oksigen pada metabolisme kondrosit
Dalam kebanyakan kes, kultur kondrosit berkembang di bawah keadaan ketegangan oksigen atmosfera. Walau bagaimanapun, diketahui umum bahawa kondrosit in vivo wujud dalam keadaan hipoksik dan ketegangan oksigen berbeza-beza di bawah pelbagai keadaan patologi. Semasa proses pematangan, perubahan ketara dalam bekalan darah ke epifisis diperhatikan. Oleh kerana vaskularisasi berbeza-beza di zon yang berbeza pada plat pertumbuhan, ketegangan oksigen di dalamnya juga berbeza-beza. C. Brighton dan R. Heppenstall (1971) menunjukkan bahawa dalam plat tibial arnab, ketegangan oksigen dalam zon hipertrofik adalah lebih rendah daripada rawan di sekelilingnya. Pengukuran beberapa parameter metabolik menunjukkan bahawa kondrosit mampu bertindak balas dengan cepat terhadap perubahan tempatan dalam kepekatan oksigen. Pertama sekali, pada ketegangan oksigen yang rendah, penggunaannya oleh kondrosit berkurangan. Dengan penurunan ketegangan oksigen daripada 21 hingga 0.04%, penggunaan glukosa meningkat, aktiviti enzim glikolitik dan sintesis asid laktik meningkat. Walaupun pada ketegangan oksigen yang rendah, jumlah mutlak ATP, ADP dan AMP kekal stabil. Data ini menunjukkan bahawa metabolisme kondrosit bertujuan untuk pemuliharaan tenaga yang maksimum. Walau bagaimanapun, aktiviti sintetik, dan oleh itu proses pembaikan, berubah di bawah keadaan hipoksik.
Ketegangan oksigen yang tinggi juga menjejaskan metabolisme kondrosit, menyebabkan penurunan dalam sintesis proteoglikan dan DNA serta degradasi matriks rawan. Kesan ini biasanya disertai dengan penghasilan radikal oksigen bebas.
Pengaruh kepekatan ion dan tekanan osmotik persekitaran terhadap fungsi kondrosit
Dalam rawan asli, kepekatan ion berbeza dengan ketara daripada tisu lain: kandungan natrium dalam medium ekstraselular ialah 250-350 mmol, dan osmolaritinya ialah 350-450 mosmol. Apabila kondrosit diasingkan daripada ECM dan diinkubasi dalam media standard (DMEM (Dulbecco's Minimal Essential Medium), osmolariti ialah 250-280.7 mosmol), persekitaran di sekeliling sel berubah secara mendadak. Di samping itu, kepekatan kalsium dan kalium dalam media standard adalah jauh lebih rendah daripada dalam tisu asli, dan kepekatan anion adalah lebih tinggi dengan ketara.
Penambahan sukrosa pada medium meningkatkan osmolaritinya dan mendorong peningkatan intraselular sementara dalam kepekatan H + dan anion kalsium dalam sitosol. Perubahan intrasel seperti itu boleh menjejaskan proses pembezaan kondrosit dan aktiviti metaboliknya. J. Urban et al. (1993) mendapati bahawa penggabungan 35 8-sulfat dan 3 H-proline oleh kondrosit terpencil yang diinkubasi dalam DMEM standard selama 2-4 jam hanyalah 10% daripadanya dalam tisu asli. Keamatan sintesis mencapai maksimum pada osmolariti medium ekstraselular 350-400 mosmol kedua-duanya dalam kondrosit yang baru diasingkan dan dalam eksplan tisu rawan. Selain itu, isipadu kondrosit meningkat sebanyak 30-40% selepas meletakkan sel terpencil dalam DMEM piawai bagi osmolariti yang ditentukan. Walau bagaimanapun, apabila mengkultur kondrosit di bawah keadaan osmolariti bukan fisiologi selama 12-16 jam, sel-sel menyesuaikan diri dengan keadaan baru, mengurangkan keamatan biosintesis berkadaran dengan peralihan dalam osmolariti persekitaran ekstraselular.
P. Borgetti et al. (1995) mengkaji kesan osmolariti medium ekstraselular terhadap pertumbuhan, morfologi, dan biosintesis kondrosit babi. Penulis menunjukkan ciri biokimia dan morfologi yang serupa bagi kondrosit yang dibiakkan dalam media dengan osmolariti 0.28 dan 0.38 mosmol. Pada osmolariti sederhana 0.48 mosmol, penurunan dalam percambahan sel dan sintesis protein diperhatikan semasa 4-6 jam pertama penanaman, tetapi parameter ini kemudiannya pulih dan akhirnya mencapai nilai kawalan. Apabila kondrosit ditanam dalam medium dengan osmolariti 0.58 mosmol, sel kehilangan keupayaan untuk mengekalkan keamatan fisiologi proses proliferatif, dan selepas 6 hari bilangan kondrosit berkurangan dengan ketara. Pada osmolariti sederhana 0.58 mosmol, perencatan mendalam sintesis protein diperhatikan. Di samping itu, apabila dibiakkan dalam media dengan osmolariti 0.28-0.38 mOsm, kondrosit mengekalkan fenotip fisiologi mereka; pada osmolariti yang lebih tinggi (0.48-0.58 mOsm), perubahan ketara dalam morfologi sel berlaku, yang ditunjukkan oleh kehilangan fenotip ciri, transformasi kondrosit menjadi sel seperti fibroblas, dan kehilangan keupayaan sel untuk memasang proteoglikan matriks. Keputusan kajian ini menunjukkan keupayaan kondrosit untuk bertindak balas terhadap turun naik terhad dalam osmolariti persekitaran ekstraselular.
Perubahan dalam kepekatan ion lain juga boleh menjejaskan proses biosintesis dalam kondrosit. Oleh itu, darjah penggabungan 35 S (sulfat) meningkat separuh dengan peningkatan kepekatan ion kalium daripada 5 mmol (kepekatan dalam medium DM EM standard) kepada 10 mmol (kepekatan dalam ECM dalam vivo). Kepekatan kalsium di bawah 0.5 mmol menggalakkan pengeluaran kolagen oleh kondrosit lembu matang, manakala kepekatan 1-2 mmol (bersamaan dengan kepekatan dalam medium DM EM standard) menyebabkan penurunan ketara dalam sintesis kolagen. Peningkatan sederhana dalam biosintesis diperhatikan pada tahap kalsium yang tinggi (2-10 mmol). Pelbagai kation mengambil bahagian dalam perlekatan kondrosit kepada protein ECM. Oleh itu, ion magnesium dan mangan menyediakan lampiran kepada fibronektin dan kolagen jenis II, manakala ion kalsium tidak mengambil bahagian dalam perlekatan kondrosit kepada protein. Oleh itu, hasil kajian yang diterangkan menunjukkan pengaruh perubahan dalam ion ekstraselular kalium, natrium, kalsium dan osmolariti medium pada fungsi biosintetik kondrosit yang diinkubasi dalam media standard.
Kesan tekanan mekanikal pada metabolisme kondrosit
Imobilisasi sendi menyebabkan atrofi rawan boleh balik, yang menunjukkan keperluan untuk rangsangan mekanikal untuk proses metabolik normal dalam ECM. Dalam kebanyakan kes, model kultur sel yang digunakan wujud di bawah tekanan atmosfera biasa. M. Wright et al. (1996) menunjukkan bahawa persekitaran mekanikal mempengaruhi metabolisme kondrosit, tindak balas sel bergantung kepada keamatan dan kekerapan pemuatan mampatan. Eksperimen dengan pemuatan pada eksplan rawan artikular utuh secara in vitro menunjukkan penurunan dalam sintesis protein dan proteoglikan di bawah tindakan pemuatan statik, manakala pemuatan dinamik merangsang proses ini. Mekanisme tepat kesan beban mekanikal pada rawan adalah kompleks dan mungkin dikaitkan dengan ubah bentuk sel, tekanan hidrostatik, tekanan osmotik, potensi elektrik dan reseptor selular permukaan untuk molekul matriks. Untuk mengkaji kesan setiap parameter ini, adalah perlu untuk mencipta sistem di mana satu parameter boleh diubah secara bebas. Sebagai contoh, kultur eksplan tidak sesuai untuk mengkaji ubah bentuk sel, tetapi ia boleh digunakan untuk mengkaji kesan umum tekanan ke atas aktiviti metabolik kondrosit. Mampatan rawan membawa kepada ubah bentuk sel dan juga disertai dengan kemunculan kecerunan tekanan hidrostatik, potensi elektrik, aliran bendalir, dan perubahan dalam parameter fizikokimia seperti kandungan air dalam matriks, ketumpatan cas elektrik, dan tahap tekanan osmotik. Ubah bentuk sel boleh dikaji menggunakan kondrosit terpencil yang direndam dalam agarose atau gel kolagen.
Beberapa sistem telah dibangunkan untuk mengkaji kesan rangsangan mekanikal pada kultur kondrosit. Sesetengah penyelidik menggunakan sistem di mana tekanan dikenakan pada kultur sel melalui fasa gas. Oleh itu, JP Veldhuijzen et al. (1979), menggunakan tekanan 13 kPa di atas atmosfera dengan frekuensi rendah (0.3 Hz) selama 15 minit, memerhatikan peningkatan dalam sintesis cAMP dan proteoglikan dan penurunan dalam sintesis DNA. R. Smith et al. (1996) menunjukkan bahawa pendedahan terputus-putus kultur kondrosit lembu primer kepada tekanan hidrostatik (10 MPa) dengan frekuensi 1 Hz selama 4 jam menyebabkan peningkatan dalam sintesis kolagen aggrecan dan jenis II, manakala tekanan malar tidak menjejaskan proses ini. Menggunakan sistem yang serupa, Wright et al. (1996) melaporkan bahawa tekanan kitaran pada kultur sel dikaitkan dengan hiperpolarisasi membran sel kondrosit dan pengaktifan saluran kalium yang bergantung kepada Ca 2+. Oleh itu, kesan tekanan kitaran dimediasi oleh saluran ion yang diaktifkan regangan dalam membran kondrosit. Tindak balas kondrosit kepada tekanan hidrostatik bergantung kepada keadaan kultur sel dan kekerapan beban yang dikenakan. Oleh itu, tekanan hidrostatik kitaran (5 MPa) mengurangkan penggabungan sulfat ke dalam monolayer kondrosit pada frekuensi 0.05, 0.25, dan 0.5 Hz, manakala pada frekuensi lebih daripada 0.5 Hz, penggabungan sulfat ke dalam eksplan tulang rawan meningkat.
M. Bushmann et al. (1992) melaporkan bahawa kondrosit dalam gel agarose mengubah biosintesis sebagai tindak balas kepada beban mekanikal statik dan dinamik dengan cara yang sama seperti organ utuh yang dikultur. Penulis mendapati bahawa beban mekanikal menjana rangsangan hiperosmotik dengan penurunan seterusnya dalam pH dalam kondrosit.
Kesan regangan mekanikal boleh dikaji pada kultur sel yang direndam dalam gel. Daya regangan boleh dicipta menggunakan vakum dikawal komputer. Apabila sistem berada di bawah tahap vakum tertentu, bahagian bawah cawan Petri dengan kultur sel dilanjutkan dengan jumlah yang diketahui, ubah bentuk adalah maksimum di tepi bahagian bawah hidangan dan minimum di tengah. Regangan juga dihantar ke kondrosit yang dikultur dalam hidangan Petri. Menggunakan kaedah ini, K. Holm-vall et al. (1995) menunjukkan bahawa dalam sel chondrosarcoma yang dikultur dalam gel kolagen (jenis II), ekspresi mRNA 2 -integrin meningkat .integrin2 β mampu mengikat kolagen jenis II. Ia dianggap sebagai mekanoreseptor, kerana ia berinteraksi dengan protein pengikat aktin, dengan itu menghubungkan ECM dan sitoskeleton.
Kesan pH pada metabolisme kondrosit
PH cecair interstisial ECM tisu rawan adalah lebih berasid daripada tisu lain. A. Maroudas (1980) menentukan pH matriks rawan artikular pada 6.9. B. Diamant et al. (1966) mendapati pH 5.5 di bawah keadaan patologi. Adalah diketahui bahawa kondrosit hidup pada PO2 yang rendah, yang menunjukkan peranan penting glikolisis (95% daripada semua metabolisme glukosa) dalam metabolisme sel-sel ini; glikolisis disertai dengan pengeluaran sejumlah besar asid laktik.
Sebagai tambahan kepada pengasidan alam sekitar oleh produk glikolisis, komponen matriks itu sendiri adalah sangat penting. Jumlah besar cas negatif tetap pada proteoglycans mengubah komposisi ionik ekstraselular: kepekatan kation bebas yang tinggi (cth, H +, Na +, K + ) dan kepekatan anion yang rendah (cth, O2, HCO3 ) diperhatikan. Di samping itu, di bawah pengaruh pemuatan mekanikal, air dikeluarkan dari ECM, yang membawa kepada peningkatan kepekatan cas negatif tetap dan tarikan lebih banyak kation ke dalam matriks. Ini disertai dengan penurunan pH persekitaran ekstraselular, yang menjejaskan pH intraselular, dengan itu mengubah suai metabolisme kondrosit. R. Wilkin dan A. Hall (1995) mengkaji kesan pH persekitaran ekstrasel dan intrasel terhadap biosintesis matriks oleh kondrosit lembu terpencil. Mereka memerhatikan pengubahsuaian dwi sintesis matriks dengan penurunan pH. Penurunan sedikit dalam pH (7.4
[ 34 ], [ 35 ], [ 36 ], [ 37 ], [ 38 ], [ 39 ], [ 40 ]
Pengaruh komposisi medium kultur pada metabolisme kondrosit
Medium untuk membiakkan kondrosit mestilah sesuai dengan keadaan eksperimen. Dalam beberapa tahun kebelakangan ini, serum anak lembu telah digunakan untuk mengoptimumkan keadaan kultur. Walau bagaimanapun, apabila menggunakan serum, beberapa perkara penting mesti diambil kira:
- pertumbuhan luar sel dari pinggir tisu dalam kultur organ,
- kebolehubahan dalam komposisi serum siri yang berbeza,
- kehadiran komponen yang tidak diketahui di dalamnya,
- peningkatan risiko gangguan dan artifak apabila mengkaji pengaruh pelbagai faktor biologi pada aktiviti metabolik sel.
Contoh yang terakhir ialah kajian tentang kesan EGF pada kondrosit rawan dalam tikus. EGF merangsang penggabungan 3 H-thymidine dan peningkatan kandungan DNA dalam budaya. Kesan ini lebih ketara pada kepekatan serum rendah (<1%), tetapi pada kepekatan tinggi (>7.5%) kesannya hilang.
Adalah diketahui umum bahawa tahap sintesis dan degradasi dalam DMEM ditambah dengan serum anak lembu meningkat dengan ketara berbanding keadaan dalam vivo. Perbezaan antara metabolisme in vivo dan in vitro mungkin disebabkan oleh perbezaan antara cecair sinovial dan medium di mana sel dikultur. Lee et al. (1997) mengkultur kondrosit lembu muda dalam agarose menggunakan medium nutrien yang mengandungi DMEM ditambah dengan 20% serum anak lembu dan sejumlah besar cecair sinovial alogenik normal. Kehadiran cecair sinovial dalam medium menyebabkan peningkatan jumlah proteoglikan, sehingga 80% daripada jumlah keseluruhan cecair sinovial. Keputusan ini menunjukkan bahawa cecair sinovial dalam kultur mendorong tahap metabolisme yang serupa dengan in vivo, dengan tahap sintesis glikosaminoglikan yang tinggi dan tahap pembahagian sel yang rendah.
G. Verbruggen et al. (1995) menunjukkan bahawa 35 S-arrpeKaHa sintesis oleh kondrosit manusia yang dibiakkan dalam agarose dalam DMEM bebas serum adalah 20-30% daripada tahap sintesis yang diperhatikan dalam DMEM ditambah dengan 10% serum anak lembu. Penulis menentukan sejauh mana IGF-1, IGF-2, TGF-R, atau insulin memulihkan pengeluaran aggrecan dalam medium bebas serum. Penulis membuat kesimpulan bahawa 100 ng/ml insulin, IGF-1, atau IGF-2 sebahagiannya memulihkan sintesis aggrecan kepada 39-53% daripada tahap kawalan. Tiada sinergi atau kumulasi diperhatikan dengan gabungan faktor yang disenaraikan. Pada masa yang sama, 10 ng/ml TGF-R dengan kehadiran 100 ng/ml insulin merangsang sintesis aggrecan kepada 90% atau lebih tahap rujukan. Akhirnya, transferin serum manusia, secara bersendirian atau digabungkan dengan insulin, tidak menjejaskan sintesis aggrecan. Apabila serum anak lembu digantikan dengan albumin serum lembu, kandungan agregat agregat menurun dengan ketara. Pengayaan medium kultur dengan insulin, IGF, atau TGF-R sebahagiannya memulihkan keupayaan sel untuk menghasilkan agregat agregat. Selain itu, IGF-1 dan insulin dapat mengekalkan homeostasis dalam kultur sel. Selepas 40 hari penanaman dalam medium yang diperkaya dengan 10-20 ng/ml IGF-1, sintesis proteoglikan dikekalkan pada tahap yang sama atau lebih tinggi berbanding dengan medium yang mengandungi 20% serum anak lembu. Proses katabolik berjalan lebih perlahan dalam medium yang diperkaya dengan IGF-1 daripada dalam medium yang diperkaya dengan larutan albumin 0.1%, tetapi agak lebih cepat dalam medium yang diperkaya dengan serum 20%. Dalam budaya tahan lama, 20 ng/ml IGF-1 mengekalkan keadaan sel yang stabil.
D. Lee et al. (1993) membandingkan kesan komposisi medium kultur (DMEM, DMEM+20% serum anak lembu, DMEM+20 ng/ml IGF-1) pada sintesis DNA dalam kultur eksplan tisu rawan, kultur monolayer, dan dalam penggantungan agarose. Apabila mengkultur dalam agarose dengan kehadiran serum, penulis memerhatikan kecenderungan untuk kondrosit berkumpul menjadi kelompok besar. Sel yang dibiakkan tanpa serum atau dengan IGF-1 mengekalkan bentuk bulat dalam agarose, dikumpulkan ke dalam kumpulan kecil, tetapi tidak membentuk agregat besar. Dalam lapisan tunggal, sintesis DNA adalah jauh lebih tinggi dalam medium yang mengandungi serum daripada dalam medium yang diperkaya dengan IGF-1; Sintesis DNA dalam yang terakhir adalah jauh lebih tinggi daripada dalam medium yang tidak diperkaya. Tiada perbezaan dalam sintesis DNA ditemui apabila kondrosit dibiakkan dalam penggantungan agarose dalam medium tidak diperkaya dan dalam medium dengan IGF-1. Pada masa yang sama, pengkulturan suspensi kondrosit dalam agarose dalam medium yang diperkaya serum disertai dengan peningkatan penggabungan radionukleotida 3 H-thymidine berbanding dengan media lain.
Vitamin C diperlukan untuk pengaktifan enzim yang terlibat dalam pembentukan struktur heliks yang stabil fibril kolagen. Kondrosit kekurangan asid askorbik mensintesis prekursor kolagen bukan heliks terhidroksilasi, yang dirembeskan secara perlahan. Pentadbiran asid askorbik (50 μg/ml) menyebabkan hidroksilasi kolagen jenis II dan IX dan rembesannya dalam kuantiti biasa. Penambahan vitamin C tidak menjejaskan tahap sintesis proteoglikan. Oleh itu, rembesan kolagen dikawal secara bebas daripada rembesan proteoglikan.