Pakar perubatan artikel itu

Penerbitan baru
Ovari
Ulasan terakhir: 04.07.2025

Semua kandungan iLive disemak secara perubatan atau fakta diperiksa untuk memastikan ketepatan faktual sebanyak mungkin.
Kami mempunyai garis panduan sumber yang ketat dan hanya memautkan ke tapak media yang bereputasi, institusi penyelidikan akademik dan, apabila mungkin, dikaji semula kajian secara medis. Perhatikan bahawa nombor dalam kurungan ([1], [2], dan lain-lain) boleh diklik pautan ke kajian ini.
Jika anda merasakan bahawa mana-mana kandungan kami tidak tepat, ketinggalan zaman, atau tidak dipersoalkan, sila pilih dan tekan Ctrl + Enter.
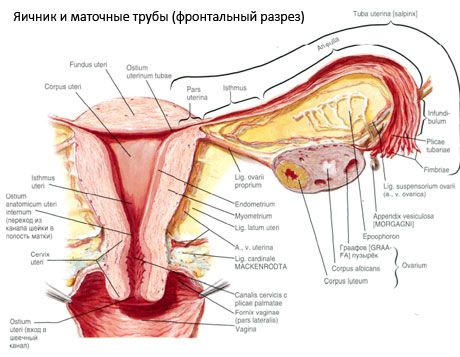
Ovari (ovarium; Greek oophoron) ialah organ berpasangan, kelenjar pembiakan wanita, terletak di rongga pelvis di belakang ligamen luas rahim. Dalam ovari, sel pembiakan wanita (telur) berkembang dan matang, dan hormon seks wanita terbentuk yang memasuki darah dan limfa. Ovari mempunyai bentuk ovoid, agak leper ke arah anteroposterior. Warna ovari adalah merah jambu. Pada permukaan ovari wanita yang telah melahirkan anak, kemurungan dan parut kelihatan - kesan ovulasi dan transformasi corpora lutea. Ovari mempunyai berat 5-8 g. Dimensi ovari ialah: panjang 2.5-5.5 cm, lebar 1.5-3.0 cm, ketebalan - sehingga 2 cm. Ovari mempunyai dua permukaan bebas: permukaan medial (facies medialis), menghadap ke rongga pelvis, sebahagiannya ditutupi oleh tiub fallopio, dan permukaan sisi (facies lateralis), bersebelahan dengan dinding sisi pelvis, dengan kemurungan yang sedikit dinyatakan - fossa ovari. Fossa ini terletak di sudut antara salur iliac luaran yang ditutup dengan peritoneum di bahagian atas dan arteri rahim dan obturator di bahagian bawah. Di belakang ovari, ureter sisi yang sepadan melepasi retroperitoneal dari atas ke bawah.
Permukaan ovari melepasi pinggir bebas (posterior) cembung (margo liber), di hadapan - ke pinggir mesenterik (margo mesovaricus), dilampirkan melalui lipatan pendek peritoneum (mesentery ovari) ke risalah posterior ligamen luas rahim. Di pinggir anterior organ ini terdapat kemurungan beralur - hilum ovari (hilum ovarii), di mana arteri dan saraf memasuki ovari, dan vena dan saluran limfa keluar. Ovari juga mempunyai dua hujung: hujung tiub atas bulat (extremitas tubaria), menghadap tiub fallopio, dan hujung rahim bawah (extremitas utenna), disambungkan ke rahim oleh ligamen ovari (lig. ovarii proprium). Ligamen ini, dalam bentuk tali bulat kira-kira 6 mm tebal, pergi dari hujung rahim ovari ke sudut sisi rahim, terletak di antara dua daun ligamen luas rahim. Alat ligamen ovari juga termasuk suspensori ligamen ovari (lig.suspensorium ovarii), iaitu lipatan peritoneum yang mengalir dari dinding pelvis kecil ke ovari dan mengandungi saluran ovari dan berkas gentian berserabut di dalamnya. Ovari diperbaiki oleh mesentery pendek (mesovarium), yang merupakan duplikasi peritoneum yang berjalan dari risalah posterior ligamen luas rahim ke pinggir mesenterik ovari. Ovari sendiri tidak dilindungi oleh peritoneum. Pinggiran ovari terbesar tiub fallopio dilekatkan pada hujung tiub ovari. Topografi ovari bergantung pada kedudukan rahim, saiznya (semasa kehamilan). Ovari adalah organ yang sangat mudah alih dari rongga pelvis kecil.
 [
[ Pembuluh dan saraf ovari
Bekalan darah ke ovari disediakan oleh aa. dan vv. ovari dan rahim. Kedua-dua arteri ovari (aa. ovaricae dextra et sinistra) berasal dari permukaan anterior aorta betul-betul di bawah arteri renal; yang kanan biasanya berasal dari aorta, dan yang kiri dari arteri renal. Menghala ke bawah dan ke sisi di sepanjang permukaan anterior otot psoas major, setiap arteri ovari melintasi ureter di hadapan (memberi cawangan kepadanya), saluran iliac luar, garis sempadan dan memasuki rongga pelvis, terletak di sini dalam ligamen suspensori ovari. Mengikuti arah medial, arteri ovari melepasi antara daun ligamen luas rahim di bawah tiub fallopio, mengeluarkan cawangan kepadanya, dan kemudian ke mesenterium ovari; ia memasuki hilum ovari.
Cabang-cabang arteri ovari secara meluas beranastomosis dengan cabang-cabang ovari arteri rahim. Aliran keluar vena dari ovari dijalankan terutamanya ke dalam plexus vena ovari, yang terletak di kawasan hilum ovari. Dari sini, aliran keluar darah melalui dua arah: melalui urat rahim dan ovari. Vena ovari kanan mempunyai injap dan mengalir ke dalam vena kava inferior. Vena ovari kiri mengalir ke dalam vena renal kiri, walaupun ia tidak mempunyai injap.
Saliran limfa dari ovari berlaku melalui saluran limfa, terutamanya dengan banyaknya di kawasan pintu organ, di mana plexus limfa subovari dibezakan. Kemudian limfa dilepaskan di sepanjang saluran limfa ovari ke nodus limfa paraaorta.
Pemuliharaan ovari
Bersimpati - disediakan oleh gentian postganglionik dari seliak (solar), plexus mesenterik dan hipogastrik yang unggul; parasimpatetik - oleh saraf sakral visceral.
Struktur ovari
Permukaan ovari ditutup dengan epitelium germinal satu lapisan. Di bawahnya terdapat lapisan protein tisu penghubung yang padat (tunica albuginea). Tisu penghubung ovari membentuk stromanya (stroma ovarii), kaya dengan gentian elastik. Bahan ovari, parenkimnya, dibahagikan kepada lapisan luar dan dalam. Lapisan dalam, terletak di tengah-tengah ovari, lebih dekat dengan pintunya, dipanggil medula (medulla ovarii). Dalam lapisan ini, dalam tisu penghubung yang longgar, terdapat banyak saluran darah dan limfa serta saraf. Lapisan luar ovari - korteks (korteks ovarii) lebih padat. Ia mengandungi banyak tisu penghubung, di mana terletak folikel ovari primer matang (folliculi ovarici primarii), folikel sekunder (vesicular) (folliculi ovarici secundarii, s.vesiculosi), serta folikel matang, folikel Graafian (folliculi ovarici maturis), serta badan kuning dan atis.
Setiap folikel mengandungi ovum pembiakan wanita, atau oosit (ovocytus). Ovum berdiameter sehingga 150 µm, bulat, mengandungi nukleus, sejumlah besar sitoplasma, yang, sebagai tambahan kepada organel selular, mengandungi kemasukan protein-lipid (kuning), glikogen, yang diperlukan untuk pemakanan ovum. Ovum biasanya menggunakan bekalan nutriennya dalam masa 12-24 jam selepas ovulasi. Jika persenyawaan tidak berlaku, ovum mati.
Telur manusia mempunyai dua selaput yang menutupinya. Di dalamnya terdapat cytolemma, iaitu membran sitoplasma telur. Di luar sitolemma adalah lapisan sel folikel yang dipanggil, yang melindungi telur dan melakukan fungsi pembentukan hormon - mereka merembeskan estrogen.
Kedudukan fisiologi rahim, tiub dan ovari disediakan oleh alat penggantung, penetapan dan sokongan yang menyatukan peritoneum, ligamen dan tisu pelvis. Alat penggantungan diwakili oleh pembentukan berpasangan, ia termasuk ligamen bulat dan luas rahim, ligamen yang betul dan ligamen penggantungan ovari. Ligamen luas rahim, ligamen ovari yang betul dan menggantung memegang rahim di kedudukan tengah. Ligamen bulat menarik fundus rahim ke hadapan dan memberikan kecondongan fisiologinya.
Alat penetapan (menambat) memastikan kedudukan rahim di tengah-tengah pelvis kecil dan menjadikannya hampir mustahil untuk beralih ke sisi, ke belakang dan ke belakang. Tetapi kerana radas ligamen berlepas dari rahim di bahagian bawahnya, kecondongan rahim ke pelbagai arah adalah mungkin. Alat penetapan termasuk ligamen yang terletak di dalam tisu longgar pelvis dan memanjang dari bahagian bawah rahim ke dinding lateral, anterior dan posterior pelvis: sacro-magic, cardinal, uterovesical dan ligamen vesikopubik.
Sebagai tambahan kepada mesovarium, ligamen ovari berikut dibezakan:
- ligamen suspensori ovari, yang sebelum ini ditetapkan sebagai ligamen infundibulopelvik. Ia adalah lipatan peritoneum dengan saluran darah (a. et v. ovari) dan saluran limfa dan saraf ovari yang melaluinya, diregangkan di antara dinding sisi pelvis, fascia lumbar (di kawasan pembahagian arteri iliac biasa ke luar dan dalam) dan hujung atas (tubal) ovari;
- ligamen ovari yang betul melepasi antara daun ligamen rahim yang luas, lebih dekat dengan daun posterior, dan menghubungkan hujung bawah ovari dengan pinggir sisi rahim. Ligamen ovari yang betul dilekatkan pada rahim antara permulaan tiub fallopio dan ligamen bulat, di belakang dan di atas yang terakhir. Dalam ketebalan pas ligamen rr. ovarii, yang merupakan cawangan terminal arteri rahim;
- ligamen apendikular-ovari Ligamen memanjang dari mesentery apendiks ke ovari kanan atau ligamen luas rahim dalam bentuk lipatan peritoneum. Ligamen tidak tetap dan diperhatikan dalam 1/2 - 1/3 wanita.
Alat sokongan diwakili oleh otot dan fascia lantai pelvis, dibahagikan kepada lapisan bawah, tengah dan atas (dalaman).
Yang paling berkuasa ialah lapisan otot atas (dalaman), diwakili oleh otot berpasangan yang mengangkat dubur. Ia terdiri daripada berkas otot yang keluar dari tulang ekor ke tulang pelvis dalam tiga arah (pubococcygeus, iliococcygeus, dan ischiococcygeus). Lapisan otot ini juga dipanggil diafragma pelvis.
Lapisan tengah otot terletak di antara tulang simfisis, kemaluan dan iskia. Lapisan tengah otot - diafragma urogenital - menduduki separuh anterior saluran keluar pelvis, yang melaluinya uretra dan vagina. Di bahagian anterior antara helaiannya adalah berkas otot yang membentuk sfinkter luar uretra, di bahagian posterior adalah berkas otot yang pergi ke arah melintang - otot melintang dalam perineum.
Lapisan bawah (luar) otot lantai pelvis terdiri daripada otot cetek, bentuknya menyerupai nombor 8. Ini termasuk bulbocavernous, ischiocavernous, sfinkter dubur luaran, dan otot perineal melintang dangkal.
Ontogenesis ovari
Proses pertumbuhan folikel dan atresia bermula pada 20 minggu kehamilan, dan pada masa penghantaran, sehingga 2 juta oosit kekal dalam ovari gadis itu. Dengan menarche, bilangan mereka berkurangan kepada 300 ribu. Sepanjang tempoh hayat pembiakan, tidak lebih daripada 500 folikel mencapai kematangan dan ovulasi. Pertumbuhan awal folikel tidak bergantung pada rangsangan FSH, terhad, dan atresia berlaku dengan cepat. Adalah dipercayai bahawa bukannya hormon steroid, peptida autokrin/parakrin tempatan adalah pengawal selia utama pertumbuhan dan atresia folikel primer. Adalah dipercayai bahawa proses pertumbuhan folikel dan atresia tidak terganggu oleh sebarang proses fisiologi. Proses ini berterusan pada semua peringkat umur, termasuk tempoh intrauterin dan menopaus, dan terganggu oleh kehamilan, ovulasi, dan anovulasi. Mekanisme yang mencetuskan pertumbuhan folikel dan bilangannya dalam setiap kitaran tertentu masih tidak jelas.
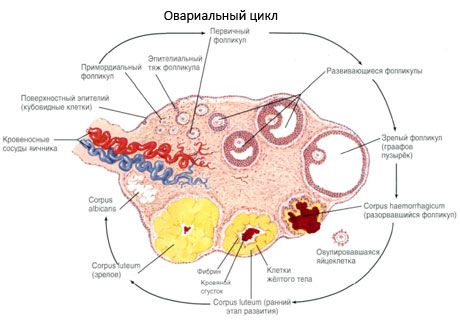
Semasa perkembangannya, folikel mengalami beberapa peringkat perkembangan. Sel germa primordial berasal dari endoderm kantung kuning telur, allantois dan berhijrah ke kawasan genital embrio pada 5-6 minggu kehamilan. Hasil daripada pembahagian mitosis yang cepat, yang berterusan dari 6-8 minggu hingga 16-20 minggu kehamilan, sehingga 6-7 juta oosit terbentuk dalam ovari embrio, dikelilingi oleh lapisan nipis sel granulosa.
Folikel preantral - oosit dikelilingi oleh membran (Zona pellucida). Sel-sel granulosa yang mengelilingi oosit mula membiak, pertumbuhannya bergantung kepada gonadotropin dan berkorelasi dengan tahap estrogen. Sel Granulosa adalah sasaran untuk FSH. Pada peringkat folikel preantral, sel granulosa mampu mensintesis tiga kelas steroid: terutamanya mendorong aktiviti aromatase, enzim utama yang menukar androgen kepada estradiol. Adalah dipercayai bahawa estradiol mampu meningkatkan bilangan reseptornya sendiri, memberikan kesan mitogenik langsung pada sel granulosa tanpa FSH. Ia dianggap sebagai faktor paracrine yang meningkatkan kesan FSH, termasuk pengaktifan proses aromatisasi.
Reseptor FSH muncul pada membran sel granulosa sebaik sahaja pertumbuhan folikel bermula. Pengurangan atau peningkatan dalam FSH membawa kepada perubahan dalam bilangan reseptornya. Tindakan FSH ini dimodulasi oleh faktor pertumbuhan. FSH bertindak melalui G-protein, sistem adenylate cyclase, walaupun steroidogenesis dalam folikel dikawal terutamanya oleh FSH, banyak faktor yang terlibat dalam proses ini: saluran ion, reseptor tyrosine kinase, sistem phospholipase utusan kedua.
Peranan androgen dalam perkembangan folikel awal adalah kompleks. Sel Granulosa mempunyai reseptor androgen. Ini bukan sahaja substrat untuk aromatisasi yang disebabkan oleh FSH kepada estrogen, tetapi juga boleh meningkatkan aromatisasi pada kepekatan rendah. Apabila tahap androgen meningkat, sel granulosa preantral lebih suka memilih bukan laluan aromatisasi kepada estrogen tetapi laluan androgen yang lebih mudah melalui 5a-reduktase untuk menukar kepada androgen yang tidak boleh ditukar kepada estrogen, dengan itu menghalang aktiviti aromatase. Proses ini juga menghalang pembentukan reseptor FSH dan LH, dengan itu menghentikan perkembangan folikel.
Proses aromatisasi, folikel dengan tahap androgen yang tinggi mengalami proses atresia. Pertumbuhan dan perkembangan folikel bergantung kepada keupayaannya untuk menukar androgen kepada estrogen.
Dengan kehadiran FSH, bahan dominan cecair folikel adalah estrogen. Dengan ketiadaan FSH - androgen. LH biasanya tiada dalam cecair folikel sehingga pertengahan kitaran. Sebaik sahaja tahap LH meningkat, aktiviti mitosis sel granulosa berkurangan, perubahan degeneratif muncul dan tahap androgen dalam folikel meningkat. Tahap steroid dalam cecair folikel lebih tinggi daripada plasma dan mencerminkan aktiviti fungsi sel ovari: sel granulosa dan theca. Jika satu-satunya sasaran untuk FSH adalah sel granulosa, maka LH mempunyai banyak sasaran - sel theca, sel stromal dan lutein dan sel granulosa. Kedua-dua sel granulosa dan theca mempunyai keupayaan untuk steroidogenesis, tetapi aktiviti aromatase mendominasi dalam sel granulosa.
Sebagai tindak balas kepada LH, sel theca menghasilkan androgen, yang kemudiannya ditukar oleh sel granulosa kepada estrogen melalui aromatisasi yang disebabkan oleh FSH.
Apabila folikel membesar, sel theca mula mengekspresikan gen untuk reseptor LH, P450 saat dan 3beta-hydroxysteroid dehydrogenase, faktor pertumbuhan seperti insulin (IGF-1) secara sinergi dengan LH meningkatkan ekspresi gen, tetapi tidak merangsang steroidogenesis.
Steroidogenesis ovari sentiasa bergantung kepada LH. Apabila folikel membesar, sel theca mengekspresikan enzim P450c17, yang membentuk androgen daripada kolesterol. Sel Granulosa tidak mempunyai enzim ini dan bergantung kepada sel theca untuk menghasilkan estrogen daripada androgen. Tidak seperti steroidogenesis, folikelogenesis bergantung kepada FSH. Apabila folikel membesar dan tahap estrogen meningkat, mekanisme maklum balas diaktifkan - pengeluaran FSH dihalang, yang seterusnya membawa kepada penurunan dalam aktiviti aromatase folikel dan, akhirnya, kepada atresia folikel melalui apoptosis (kematian sel terprogram).
Mekanisme maklum balas estrogen dan FSH menghalang perkembangan folikel yang telah mula berkembang, tetapi bukan folikel dominan. Folikel dominan mengandungi lebih banyak reseptor FSH, yang menyokong percambahan sel granulosa dan aromatisasi androgen kepada estrogen. Di samping itu, laluan paracrine dan autokrin bertindak sebagai penyelaras penting perkembangan folikel antral.
Pengatur autokrin/paracrine terdiri daripada peptida (inhibin, activin, follistatin), yang disintesis oleh sel granulosa sebagai tindak balas kepada FSH dan memasuki cecair folikel. Inhibin mengurangkan rembesan FSH; activin merangsang pembebasan FSH dari kelenjar pituitari dan meningkatkan tindakan FSH dalam ovari; follistatin menyekat aktiviti FSH, mungkin dengan mengikat aktivin. Selepas ovulasi dan perkembangan korpus luteum, inhibin berada di bawah kawalan LH.
Pertumbuhan dan pembezaan sel ovari dipengaruhi oleh faktor pertumbuhan seperti insulin (IGE). IGF-1 bertindak ke atas sel granulosa, menyebabkan peningkatan dalam kitaran adenosin monofosfat (cAMP), progesteron, oksitosin, proteoglycan, dan inhibin.
IGF-1 bertindak pada sel theca, menyebabkan peningkatan pengeluaran androgen. Sel theca pula menghasilkan faktor nekrosis tumor (TNF) dan faktor pertumbuhan epidermis (EGF), yang juga dikawal oleh FSH.
EGF merangsang percambahan sel granulosa. IGF-2 ialah faktor pertumbuhan utama dalam cecair folikel, dan IGF-1, TNF-a, TNF-3, dan EGF juga terdapat di dalamnya.
Gangguan peraturan paracrine dan/atau autokrin terhadap fungsi ovari kelihatan memainkan peranan dalam gangguan ovulasi dan dalam perkembangan ovari polikistik.
Apabila folikel antral berkembang, kandungan estrogen dalam cecair folikel meningkat. Pada puncak peningkatannya, reseptor untuk LH muncul pada sel granulosa, luteinisasi sel granulosa berlaku, dan pengeluaran progesteron meningkat. Oleh itu, dalam tempoh praovulasi, peningkatan dalam pengeluaran estrogen menyebabkan kemunculan reseptor LH, LH, seterusnya, menyebabkan luteinisasi sel granulosa dan pengeluaran progesteron. Peningkatan progesteron mengurangkan tahap estrogen, yang nampaknya menyebabkan puncak kedua FSH di tengah kitaran.
Ovulasi dianggap berlaku 10-12 jam selepas puncak LH dan 24-36 jam selepas puncak estradiol. LH dianggap merangsang pembahagian pengurangan oosit, luteinisasi sel granulosa, dan sintesis progesteron dan prostaglandin dalam folikel.
Progesteron meningkatkan aktiviti enzim proteolitik, yang bersama-sama dengan prostaglandin mengambil bahagian dalam pecah dinding folikel. Puncak FSH yang disebabkan oleh progesteron membolehkan oosit keluar dari folikel dengan menukar plasminogen kepada plasmin enzim proteolitik, dan menyediakan bilangan reseptor LH yang mencukupi untuk perkembangan normal fasa luteal.
Dalam masa 3 hari selepas ovulasi, sel granulosa meningkat dalam saiz, dan vakuol ciri yang dipenuhi dengan pigmen, lutein, muncul di dalamnya. Sel theca-luteal membezakan daripada theca dan stroma dan menjadi sebahagian daripada corpus luteum. Kapilari yang menembusi korpus luteum berkembang sangat cepat di bawah pengaruh faktor angiogenesis, dan dengan vaskularisasi yang lebih baik, pengeluaran progesteron dan estrogen meningkat. Aktiviti steroidogenesis dan jangka hayat korpus luteum ditentukan oleh tahap LH. Korpus luteum bukanlah pembentukan selular yang homogen. Sebagai tambahan kepada 2 jenis sel luteal, ia mengandungi sel endothelial, makrofaj, fibroblas, dll. Sel luteal yang besar menghasilkan peptida (relaxin, oxytocin) dan lebih aktif dalam steroidogenesis dengan aktiviti aromatase yang lebih besar dan sintesis progesteron yang lebih besar daripada sel kecil.
Puncak progesteron berlaku pada hari ke-8 selepas puncak LH. Adalah diperhatikan bahawa progesteron dan estradiol dirembeskan secara episod dalam fasa luteal dalam korelasi dengan output pulsatil LH. Dengan pembentukan korpus luteum, kawalan pengeluaran inhibin berpindah dari FSH ke LH. Inhibin meningkat dengan peningkatan estradiol sebelum puncak LH dan terus meningkat selepas puncak LH, walaupun tahap estrogen menurun. Walaupun inhibin dan estradiol dirembes oleh sel granulosa, mereka dikawal oleh laluan yang berbeza. Penurunan inhibin pada akhir fasa luteal menyumbang kepada peningkatan FSH untuk kitaran seterusnya.
Korpus luteum berkurangan dengan cepat - pada hari ke-9-11 selepas ovulasi.
Mekanisme degenerasi tidak jelas dan tidak berkaitan dengan peranan luteolitik estrogen atau mekanisme yang berkaitan dengan reseptor, seperti yang diperhatikan dalam endometrium. Terdapat penjelasan lain untuk peranan estrogen yang dihasilkan oleh korpus luteum. Adalah diketahui bahawa estrogen diperlukan untuk sintesis reseptor progesteron dalam endometrium. Estrogen fasa luteal mungkin diperlukan untuk perubahan berkaitan progesteron dalam endometrium selepas ovulasi. Perkembangan reseptor progesteron yang tidak mencukupi, akibat tahap estrogen yang tidak mencukupi, mungkin merupakan mekanisme tambahan untuk ketidaksuburan dan kehilangan kehamilan awal, satu lagi bentuk kekurangan fasa luteal. Adalah dipercayai bahawa jangka hayat korpus luteum ditubuhkan pada masa ovulasi. Dan ia pasti akan mundur jika ia tidak disokong oleh human chorionic gonadotropin akibat kehamilan. Oleh itu, regresi korpus luteum membawa kepada penurunan tahap estradiol, progesteron dan inhibin. Pengurangan dalam inhibin menghilangkan kesan penindasannya pada FSH; penurunan dalam estradiol dan progesteron membolehkan pemulihan rembesan GnRH yang sangat cepat dan penyingkiran mekanisme maklum balas daripada kelenjar pituitari. Pengurangan dalam inhibin dan estradiol, bersama-sama dengan peningkatan dalam GnRH, menimbulkan FSH berbanding LH. Peningkatan FSH membawa kepada pertumbuhan folikel dengan pemilihan folikel dominan yang seterusnya, dan kitaran baru bermula jika kehamilan tidak berlaku. Hormon steroid memainkan peranan utama dalam biologi pembiakan dan fisiologi am. Mereka menentukan fenotip manusia, mempengaruhi sistem kardiovaskular, metabolisme tulang, kulit, kesejahteraan umum dan memainkan peranan penting dalam kehamilan. Tindakan hormon steroid mencerminkan mekanisme intrasel dan genetik yang diperlukan untuk menghantar isyarat ekstraselular kepada nukleus sel untuk menyebabkan tindak balas fisiologi.
Estrogen meresap menembusi membran sel dan mengikat reseptor yang terletak di dalam nukleus sel. Kompleks reseptor-steroid kemudian mengikat DNA. Dalam sel sasaran, interaksi ini membawa kepada ekspresi gen, sintesis protein, dan fungsi sel dan tisu tertentu.